What is Mycorrhizal Applications?
As the world’s leading manufacturer and supplier of mycorrhizal inoculant products and ingredients, Mycorrhizal Applications (MA) researches, produces, and markets mycorrhizal fungi which accelerate plant vigor by greatly increasing the surface absorbing area of roots, producing a healthier root system. These specialized fungi colonize plant roots to create a symbiotic root-and-mycelial network within the surrounding soils, increasing efficiency in nutrient and water absorption to optimize plant performance. MA’s MycoApply® line of mycorrhizal inoculants is utilized by landscapers, farmers, forest nurseries, restoration & erosion control specialists, greenhouses, nurseries, soil media manufacturers, and professional horticulturalists to maximize plant success and improve return on investment.
What Are Mycorrhizae?
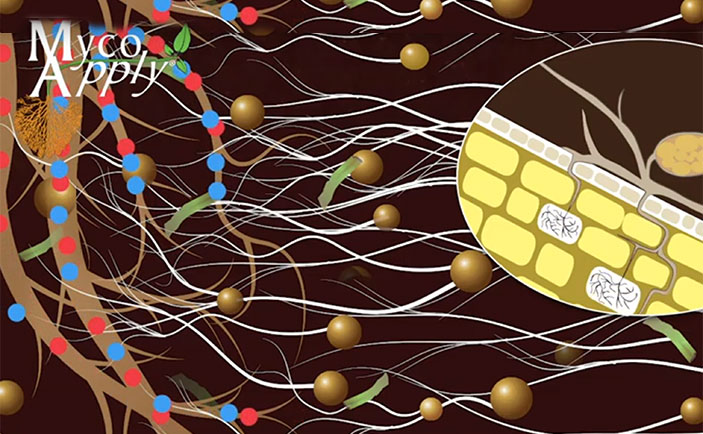
“Myco” – “rhiza” literally means “fungus” – “root” and describes the mutually beneficial relationship between the plant and root fungus (mycorrhizae is the plural form of mycorrhiza). These specialized fungi colonize plant roots and extend far into the soil. Mycorrhizal fungal filaments in the soil are truly extensions of root systems and are more effective in nutrient and water absorption than the roots themselves. Approximately 95 percent of plant species on the planet form a symbiotic relationship with the beneficial mycorrhizal fungi.
Are They Important?
Mycorrhizal fungi increase the surface absorbing area of roots, thereby greatly improving the ability of the plant to access soil resources. Mycorrhizae are an essential part of a healthy soil microbiome, and several miles of fungal filaments can be present in less than a thimbleful of soil. Mycorrhizal fungi increase nutrient uptake not only by increasing the surface absorbing area of the roots, but also release powerful organic compounds into the soil that help to solubilize hard-to-capture nutrients, such as organic nitrogen, phosphorus, iron and other “tightly bound” soil nutrients. This extraction process is particularly important in plant nutrition and explains why non-mycorrhizal plants require high levels of fertilization to maintain their health. Mycorrhizal fungi form an intricate web that captures and assimilates nutrients, conserving the nutrient capital in soils.
Do You Need Them?
Undisturbed soils are full of beneficial soil organisms including mycorrhizal fungi. Research indicates, however, many common practices can degrade the mycorrhizae-forming potential of soil. Tillage, removal of topsoil, erosion, site preparation, compaction, fumigation, invasion of weeds, and leaving soils fallow are some of the activities that can reduce or eliminate these beneficial soil fungi. Scientific studies indicate mycorrhizal populations are slow to recolonize naturally, therefore, reintroducing mycorrhizal fungi in areas where they have been lost or in artificial growing media can dramatically improve plant performance with less water and fertilizer and at a reduced cost.